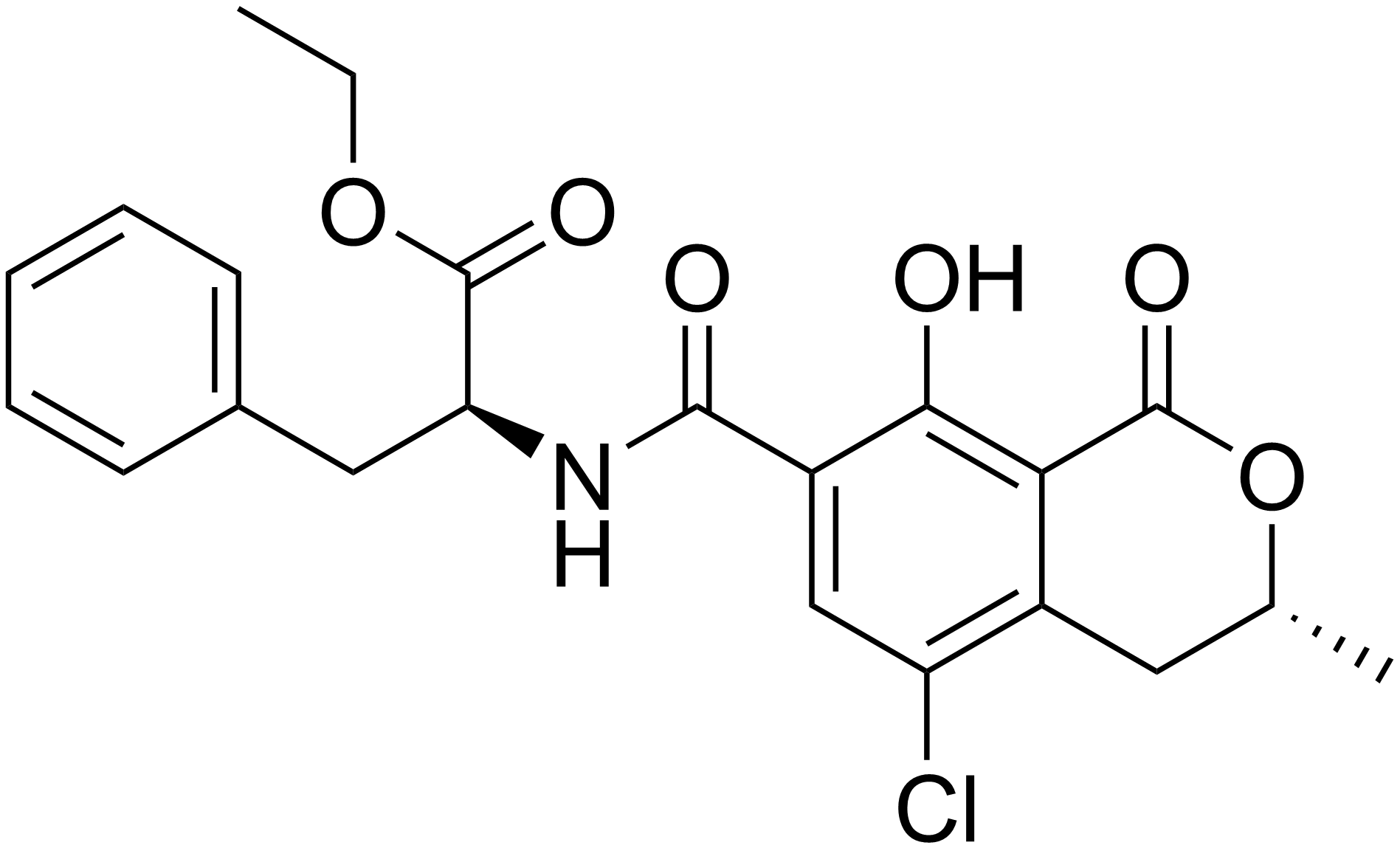
Okratoksin A
Suda az, polar organik çözücüler ile seyreltik sulu sodyum bikarbonat çözeltisinde iyi çözünen, renksiz ve kristal yapıda bir bileşiktir. UV ışınları altında mavi renkte floresan verir. Okratoksin A, 7-karboksi-5-kloro-8- hidroksi-3,4 dihidro-3R-metil-izokumarin içerir ve L-fenilalanine karboksi grup üzerinden bağlanır.
Kimyasal yapısında fenilalanin, Cl ve OH içeren dihidroizokumarin bulunur. Okratoksin A'nın Cl içermeyen türevi Okratoksin B, etilester türevi ise okratoksin C'dir. Okratoksin B ve okratoksin C gıdalarda görülseler de düşük konsantrasyonda bulunduklarından fazlaca önem taşımazlar. Okratoksin A, pankreas ve ince bağırsakta α-kimotripsin ve karboksipeptidaz enzimleri ile okratoksin α’ya hidroliz edilmektedir.
Okratoksin A büyük miktarlarda üretilir ve sık sık gıda ve yemlerde doğal kontaminant olarak rastlanır. Farklı gıdalarda yapılan çalışmalar okratoksin A’nın sıcaklığa karşı dayanıklı olduğunu göstermiştir. Patterson (1977), Josefson ve Möller (1980), Scott (1984), domuz eti ve sakatatların 150-160 ºC sıcaklıkta pişirilmesiyle toksin miktarında % 20 azalma meydana gelirken, yağ dokusunda bulunan toksinde hiç kayıp olmadığını bildirmektedir. Okratoksinle kontamine olmuş un ile yapılan ekmekte pişirme işlemi neticesinde az bir kaybın olduğu gözlenmişken, bisküvi yapımında çok daha fazla toksinin parçalandığı bildirilmiştir (Metin 2006).
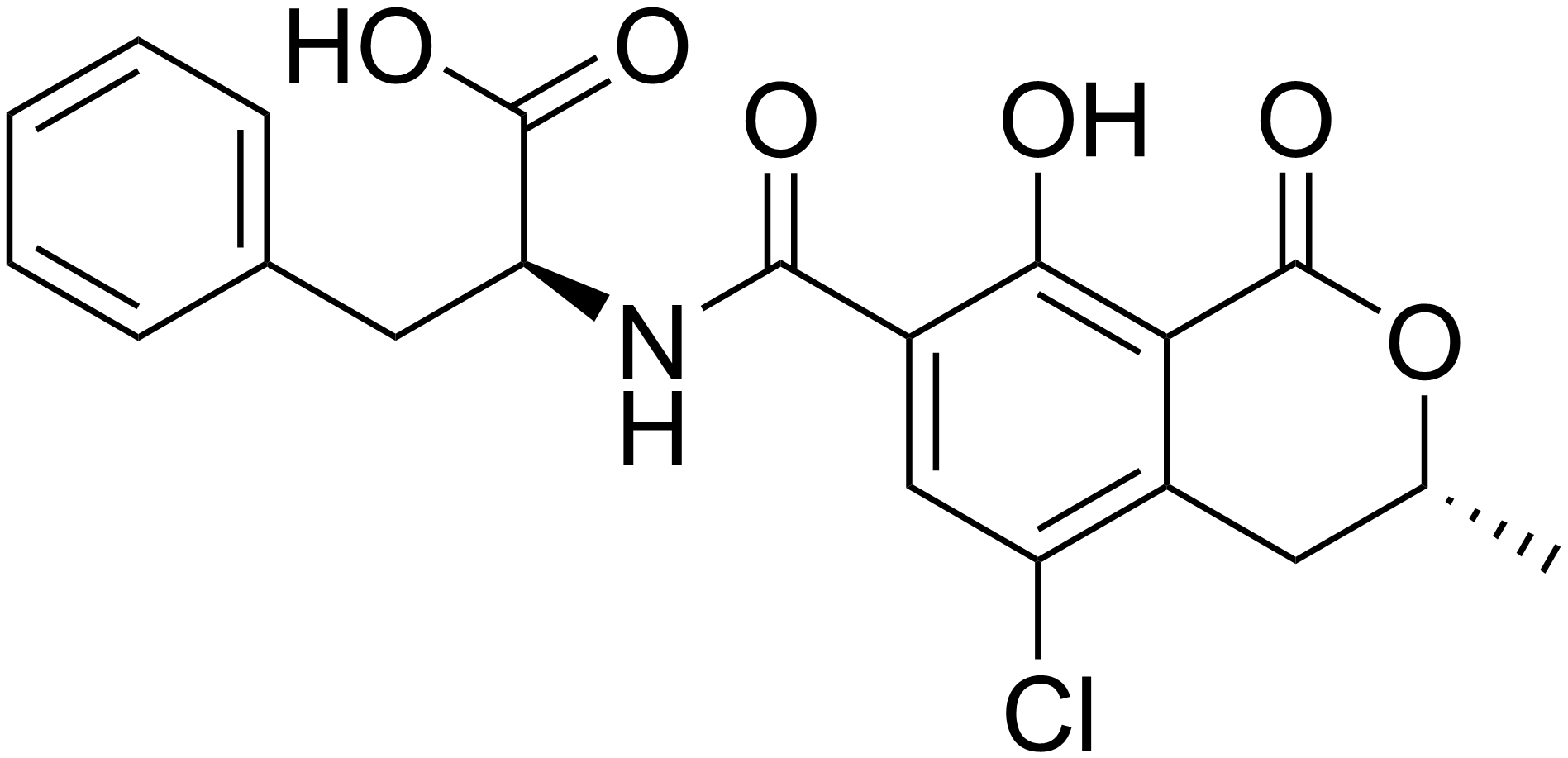
Okratoksin B
Okratoksin B (OTB) A. ochraceus tarafından üretilen sekonder bir metabolittir.
OTA ile birlikte oluşabilir. OTB, moleküler yapının 5. pozisyonundaki klorun eksikliği nedeni ile OTA’dan ayrılmaktadır. Yapısal olarak OTA’ya çok benzer olmasına rağmen daha az toksisiteye sahip olduğu düşünülmektedir. OTB’nin OTA’ya göre daha hızlı metabolize olduğu rapor edilmiştir ( Mally vd 2005).
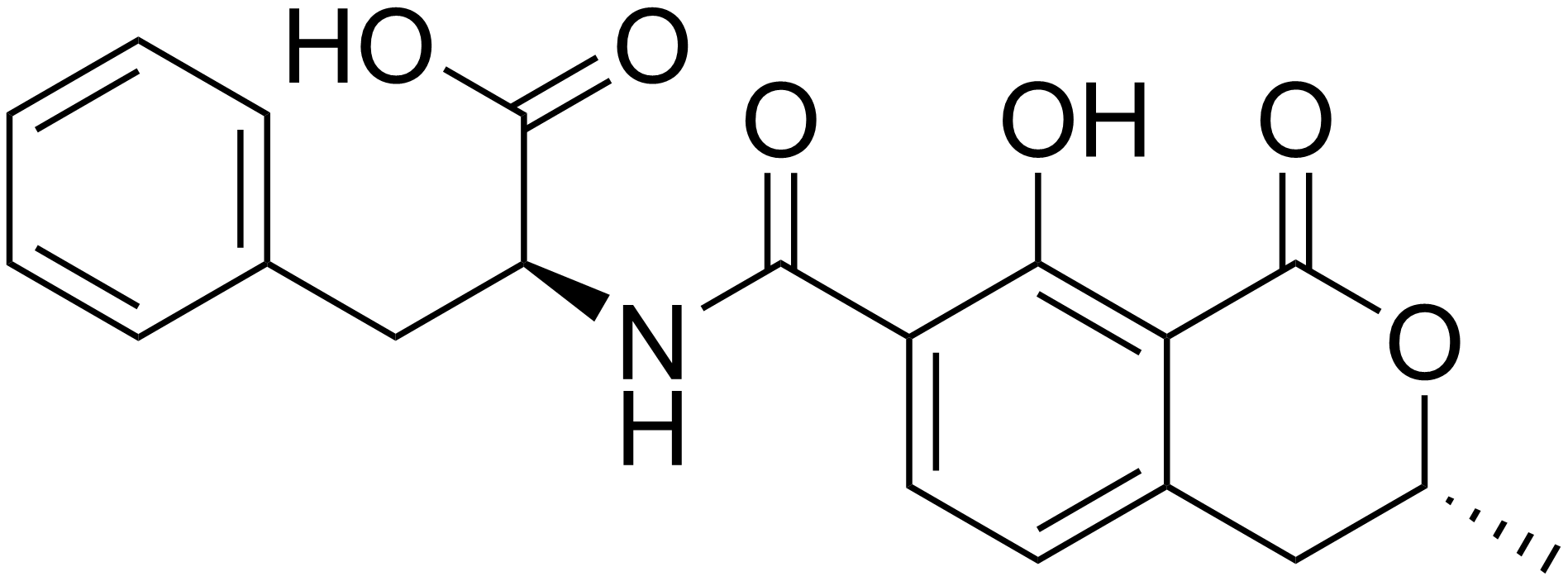
Okratoksin C
OTA’nın etil esteri olan Okratoksin C’ye (OTC) gıda ve hayvan yemlerinde doğal kontaminant olarak ender rastlanır. Ancak çok yüksek konsantrasyonda OTA içeren yemlerde OTC varlığına rastlanmaktadır. Galtier ve Alvinerie (1976) tarafından yapılan bir çalışmada inek ve koyunlara ait işkembe sıvılarının OTA’yı hidrolize ettiği ve OTC’ye dönüştürdüğü tespit edilmiş ve araştırmacılar bu durumun geviş getiren hayvanların ön midelerinde bulunan protozoa ve bakteriyel enzimler tarafından gerçekleştirildiğini ileri sürmüşlerdir. Bu nedenle hayvansal orijinli gıdalarda kalıntı olarak bulunan OTC’nin halk sağlığı problemlerine yol açabileceği sonucu ortaya çıkmaktadır (Fuchs vd 1984).
Okratoksin A çoğunlukla subtropikal ve ılıman iklimlerdeki tahıllarda, kahve çekirdekleri, kakao, kuru incir, yaş ve kuru üzüm, fındık, yer fıstığı ve karışık yemlerde sıkça saptanan en önemli mikotoksinlerden biri olarak kabul edilmektedir. Ayrıca bira, şarap ve üzüm suyu gibi yan ürünlerde de varlığı tespit edilmiştir.
Bitkisel ürünler içerisinde buğday, arpa, yulaf, çavdar, mısır ve pirinç gibi tahıllar OTA'yı yüksek konsantrasyonda içermelerinden dolayı, hem insanlar hem de hayvanlar açısından en önemli kontaminasyon kaynağıdır (Tunail 2001).
Gıda ve Tarım Organizasyonu (FAO) tarafından OTA üzerinden belirlenen Avrupa Birliği tarafından ise bebek mamaları için müsaade edilen düzey 1 µg/kg; tahıllar için ise 5 µg/kg olarak belirlenmiştir (Girgin vd 2001). İnsanlarda OTA için tolere edilebilir günlük alım düzeyi (Tolerable Daily IntakeTDI) Dünya Sağlık Örgütü tarafından 14 ng/kg olarak bildirilmiştir. Gıda Toksikolojisi ve Risk Değerlendirmesi Nordik Çalışma Grubu tarafından yapılan değerlendirmede kabul edilen TDI 5 ng/kg’dır. Kanada Uzmanlar Grubu ise TDI için 1.2-5.7 ng/kg değerini önermektedir (Barlow vd 2008).
Farklı gıda maddelerindeki izin verilen Okratoksin A limitleri
| Ürün | Limit (µg/kg) |
| Çocuk ve bebek mamaları | 0.5 - 5 |
| Yiyecekler | 2 - 50 |
| Hayvan yemi | 5 - 300 |
| Şarap | 0.2 - 1 |
| Bira | 0.2 |
| Yeşil kahve tohumu | 8 |
| Kavrulmuş kahve ve ürünleri | 4 |